
Muhammad Aslam ( Department of Physiology, Army Medical College, Rawalpindi. )
Muhammad Amjad Hameed ( Department of Physiology, Army Medical College, Rawalpindi. )
September 1989, Volume 39, Issue 9
Review Articles
The polyamines (putrescine, spermidine and spermine) are a group of small basic molecules distributed universally in all the living organisms including animals, plants and viruses1. Two polyamine derivatives, putreanine and spermic acid, probably metabolites of sperniidine and spermine respectively, have also been isolated from nammalian nervous system. 2 It was not until 1968 that the enzymology of polyamines in nammalian tissues began to be unrevealed. The enzymes needed for the synthesis of polyamines are: (1) Ornithine decarboxylase -catalysing the decarboxylation of L-ornithine to yield putrescine (2) S-adenosyl methionine decarboxylase - forming decarboxylated adenosyhnethionine from adenosylmethionine. (3)Spermidine synthetase - which transfers the polymine group from decarboxylated adenosylmethionine to putrescine to yield spernildine, methyl thioa-denosines are formed as other reaction products. (4) spermine synthetase - catalyses the formation of spermine; the reaction is analogous to that of spermidine synthetase (Figure -1).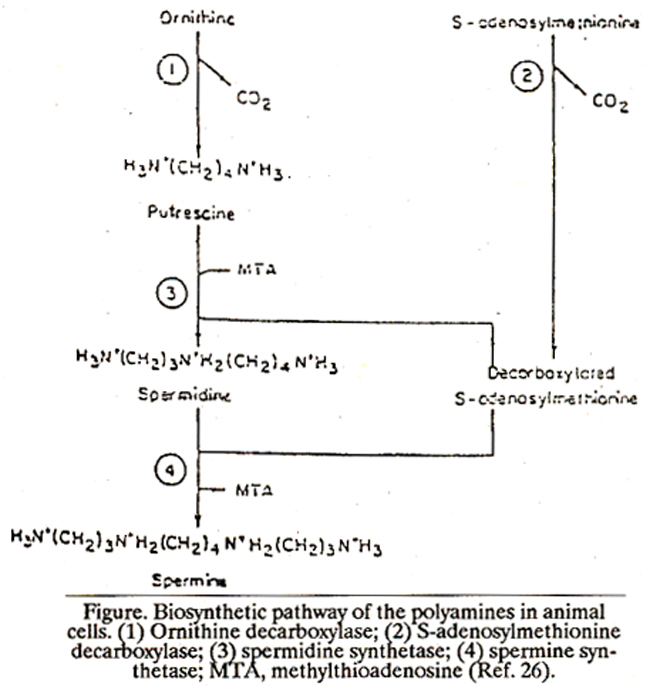
Relatively little is known about the catabolism and excretion of polyamines in animals tissues. It has been reported that about 40% of polyamines in the rat which are eliminated, are excreted in the urine as or acetylated derivatives and about 60% are catabolised by the oxidases. 3 The enzymes involved in the specific biosynthetic steps of polyamine production have been purified and characterised from a variety of animal sources. All of these appear to be soluble cytosol enzymes and their general properties are relatively well- documented. Ornithine decarboxylase (ODC) provides the biosynthetic route giving rise to putrescine in a reaction that maybe rate-limiting for polyamine synthesis. Polyamines and ODC activity is normally very low in quiscent tissues, but is significantly higher in rapidly proliferating cells. A large number of stimuli4 cause a rapid increase (100 to 200 fold) in the level of ODC activity, both in vivo and in cultured mammalian cells. Some of the most effective stimuli are malignant growth, high amino- acid intake, embryonic development, renal hypertrophy, cardiac enlargement, liver regeneration after hepatectomy, virus infection, thiocetamide administration, epidermal growth factor, exposure to cold and other stresses and administration of various hormones. The exact mechanism of control of ODC is not fully known but results from various studies5 suggest that induction of ODC in many tissues involves cyclic AMP and guanine nucleotides and its inhibition, the antizyme.
HORMONAL CONTROL OF POLYAMINES
Most attention has focussed on the role of polyamines in normal growth and differentiation, though some studies have proposed that they may mediate the responses to hormonal stimuli in peripheral target organs.
Luteinising Hormones-releasing hormone
In recent studies6,7, luteinising hormones-releasing hormone (LHRH) analogue has been reported to reduce oestrogen-induced ODC activity in renal and uterine tissues. Whereas, no change in pituitary ODC activity was found after an injection of LHRH in ovariectomised rats. 8 Very recently, LHRH has been postulated to induce activity of the enzyme in vitro9 contributing for preovulatory surge.
Corticotrophin Releasing Hormone
It has been shown10 that stimulation of pituitary gland with corticotrophin releasing activity causes an increase in the polyamine content of the tissue and that putrescine can stimulate the release of adrenocorticotrophic hormone. 11
Growth Hormone
A rapid decrease in the concentration of hepatic spermidine was observed after hypophysectomyofthe rat12 suggesting that the metabolism of mammalian polyamines might be controlled by growth hormone. The content of liver spermidine was restored to the normal levels by treatment with growth hormone possibly due to stimulation of ODC activity. The dramatic stimulation of ODC activity in liver, kidney and heart by growth hormone, either in normal or in hypophysectomised rats, has been confirmed. 13
Thyroid Stimulating Hormone
A preferential role for spermidine in stimulation of thyroid hormone output from thyroid gland in the guinea pig has been reported. 14 The biochemical event mediated by thyroid stimulating hormone in thyroidal follicular cells is the induction of ODC activity. Furthermore, thyroxine itself has been reported to stimulate ODC activity in the rat.
Adrenocorticotrophic Hormone
Adrino-corticotrophie hormone (ACTH) has been found to cause an early stimulation of ODC activity in the adrenal cortex of hypophysectomised rats. The stimulation of ODC by ACTH is preceded by an increase in cyclic AMP suggesting a quantitative relationship between cyclic AMP and ODC activity in the adrenal gland. 15 Recently, it has been suggested that polyamines mayfunction as intracellular messengers for some of the ACTH effects in bovine adrenocortical cells. 16
Follicle Stimulating Hormone
There have been a few studies showing involvement of polyamines in follicle stimulating hormone (FSH) secretion from pituitaries in the rat. A decrease of in vivo release of pituitary FSH in intact and is castrated, testosterone-treated rats in response to an injection of naturally occurring polyamine (especially putrescine) isolated from porcine hypothalamus was observed. 17 Whereas, spermidine has been reported to enhance FSH release from pituitaries incubated in vitro. It has been demonstrated that FSH stimulates ODC activity in the ovary and testes. 18 Recently, it has been suggested that the polyamines (especially spermine) biosynthetic pathways of rat Sertoli cells appears to be regulated by FSH.
Luteinising Hormone
In mammalian species, luteinising hormone (LH) has been shown to be a potent activator of ovarian ODC activity In vivo and in vitro19. The activity of ovarian ODC can also be stimulated by human chorionic gonadotrophin in immature rats. In the laying hen, it has been found that bovine LH can stimulate ODC activity in granulosa cells in vito20 Very recently21 putrescine has been reported to play a permissive role inthe generation of prevulatory surge of LH and the subsequent ovulation.
Prolactin
Prolactin has been shown to stimulate ODC activity in adrenal gland, thymus and mammary tumors. 22 However, experiments of Persson and others8 led them to conclude that an increase in polyamine biosynthesis in the anterior pituitary gland is not related to the secretion of prolactin as administration of throtrophin-releasing hormone (which causes prolactin release) failed to stimulate pituitary ODC activity in rats.
Ovarian Steroids
Injection of oestrogens into immature rats resulted in a marked increase in ODC activity of uterine homogenates. Progesterone has also been reported to stimulate the activity of ODC in the chick oviducts. Moreover, this steroid has also been shown to increase ODC activity and subsequent polyamine concentration in the guinea pig uterus and in rat kidney cells. 23 Recently, it has been suggested that oestradiol and progesterone act synergistically to enhance ODC activity andputrescine concentration in the anterior pituitary gland which, in turn, leads the culmination of preovulatory surge and subsequent ovulation. 24-26.These studies suggest that ODC activity and polyamines can be altered in target organs exposed to specific hormone in vitro and in vivo. If the role of polyamines is established as a second or third messenger for hormonal action at cellular level, the venue of mechanistic endocrinology would change. Furthermore, the possibility of the role of putrescine to induce fertility in an ovulatory condition due to lack of LH surge cannot be ruled out in future.
REFERENCES
1. Tabor, H. and Tabor, C.W. Spermidine, spermine and related amines. Pharmacol. Rev., 1964; 16:245.
2. Imaoka,N. and Matsuoka, Y. Isolation and identification of spermic acid, N. N, his (2 carboxyethyl)-1,4-diaminobutane from bovine brain. J. Neurochent, 1974; 22:859.
3. Seiler, N., Bokenivs, F.N. and Knodgen, B. The influence of catabolic reactions on polyamine excretion. Biochem. 3., 1985; 208:189.
4. Pegg. A.E., Seely, J.E., Poso, H., Ragione, ED. and Zagon, 1.5. Polyamine biosynthesis and interconversion in rodent tissues. Fed. Proc., 1982; 41:305.
5. Wright, J.M. and Boyle, S.M. Negative control of ornithine decarboxylase and arginine decarboxylase by adenosine-3:5 cyclic monophosphate in Escherichia Coli. Mol. Gen. Genet., 1982; 186:482.
6. Prasad, M.S., Vani, V.V., Raju, RS. and Reddy, 1CR. Inhibition of sex-steroid hormone-induced ornithine decarboxylase and poly (A) polymerase activities by a GnRH agonist in the rat kidney. 3. Steroid Biochem; 1985; 23:793.
7. Rao, I.M. and Rèddy, P.R. Direct inhibitory effect of gonadotropin releasing hormone in the uterus of rat. Life Sci., 1984; 34:2257.
8. Persson, L., Nilsson, M. and Rosengren, L.E. Ornithine decarboxylase activity and polyamines in the anterior pituitary gland during the rat oestrous cycle. J.Endocrinol., 1985; 107:83.
9. Aslam, M., Nicholson, S. Gillham, B. and Jones, M. Permissive role of ornithine decarboxylase and putrescine in the luteinising hormone surge. Neuroendocrinology, 1987; 45:473.
10. Morgan-Jones, LM., Gillham, B., Aslam, M. and Jones, M.T. Polyamines and anterior pituitary gland hormone secretion. Proc. Inaug. Meeting Oxford. Br. Neuroendoer. Cr., 1985: Abs. 44.
11. Nicholson, S.A. Further studies on recovery of the responsiveness of the pituitary gland after chronic prednisolone treatment in the rat. 3. Physiol., 1986; 327:63P.
12. Kostyo, J.L Changes in polyamine content of rat liver following hypophysectomy and treatment with growth hormone. Biochem. Biophys. Res. Commun., 1966;23:150.
13. Antony, P., Gibson, K.!. and Hanis, D.Ornithine decar boxylasein the isolated perfused rat heart. Biochem. Soc. Trans., 1975; 3:272.
14. Corcoran, A.E. and Smyth, P.P.A. Polyamines are módiators of thyroid stimulation. 3. Endocrinol., 1986;108(S):abs.178.
15. Richman, R., Dobbins, C., Voina, S., Underwood, L., Mahafee, D., Gitelman, IIJ., Van Wyk, J. and Ney, RL Regulation of adrenal ornithine decarboxylase by adrenocorticotropic hormone and cyclic AMP. 3. Clin. Invest., 1973; 52:2007.
16. Feige, JJ., Madani, C. and Chambaz, E.A. Hormonal control of polyamine levels in bovine adrenocortical cells. Endocrinology, 1986; 118:1059.
17. White, W.F., Cohen, A.A., Rippel, RH. and Story, J.E. Some hypothalamic polyamines that deplete pituitary follicle stimulating hormone. Endocrinology, 1968;82:741.
18. Rani, C.L.S. and Moudgal, N.R. Examination of the role of FSH in oestrogen biosynthesis In vivo and in vitro in the ovary of cyclic hamster. Steroids, 1978; 32:435.
19. Kay, A.M., Leekson, I., Lamprecht, A., Gruss, It, Tsafriri, A and Linder, H.R Stimulation of ornithine decarboxylase activity by LII in immature and adult rat ovaries. Biochemistry, 1973; 12:3072.
20. Armstrong, D.G. Ornithine decarboxylase activity in small ovarian follicles from the laying hen (Callus deomesticus). J. Endocr., 1987; 112:183.
21. Aslam, M., Nicholson, S.A., Oon, B.B., Gillham, B. and Jones, M.T. A role for ornithine decarboxylase in the pro-oestrous LII surge in rats. Proc. 1st. Int. Cong. Neuroendocr., 1986; San Franciso (USA); p.24 Abs.6.
22. Frazier, RP. and Coslow, M.E. Prolactin stimulation of ornitbine decarboxylase activity in cultured rat mammary tumours. Exp. Cell Res., 1982; 138:39.
23. Lin, Y.E., Loring, J.M. and Villee, C.A. Diethylstil bestrol stimulated ornithine decarboxylase in kidneycells in culture. Biochem. Biophys. Res. Commun., 1980; 1393.
24. Aslam, M., Nicholson, S.A., Gillham,B. and Gones, M.T. Regulation of ornithine decarboxylase activity in the rat pituitary by ovarian steroids. 3. Endocr., 1987; 112 (5): Abs. 71.
25. Nicholson, S.A., Aslam, M., Chuang, T.T., Gillham, B. and Jones, M.T. Effect of difluoromethylornithine on the LII surge and subsequent ovulation in the rat. J. Endocr., 1988; 117:447.
26. Goyns, M.II. The role of polyamines in animal cell physiology. J. Theor. Biol., 1982; 32:57.
Journal of the Pakistan Medical Association has agreed to receive and publish manuscripts in accordance with the principles of the following committees: